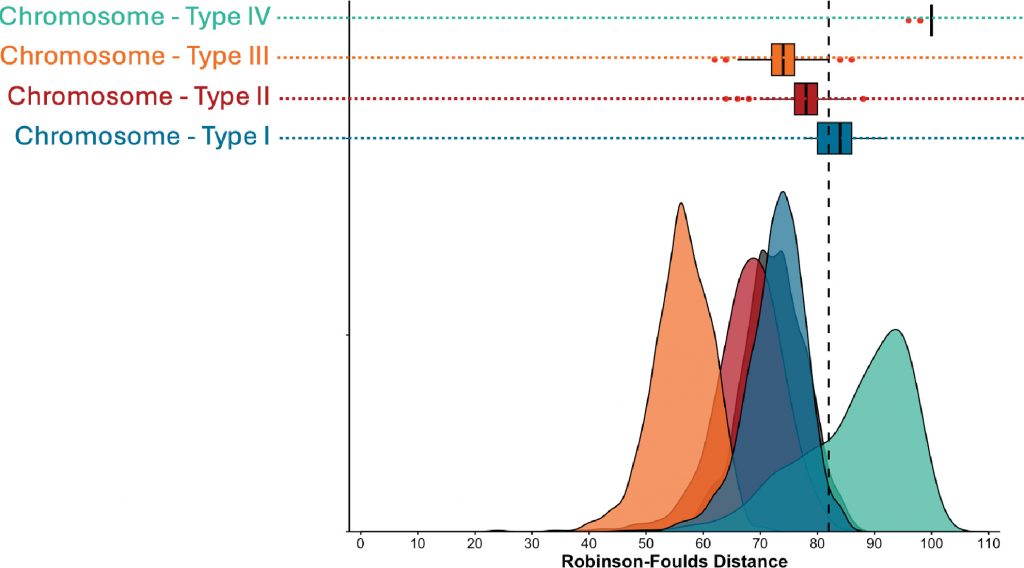
Our undergraduates are AWESOME!
Amartya, Dave, and Julienne presented their hard work at URS and IGB. Look at those proud mentors 🙂
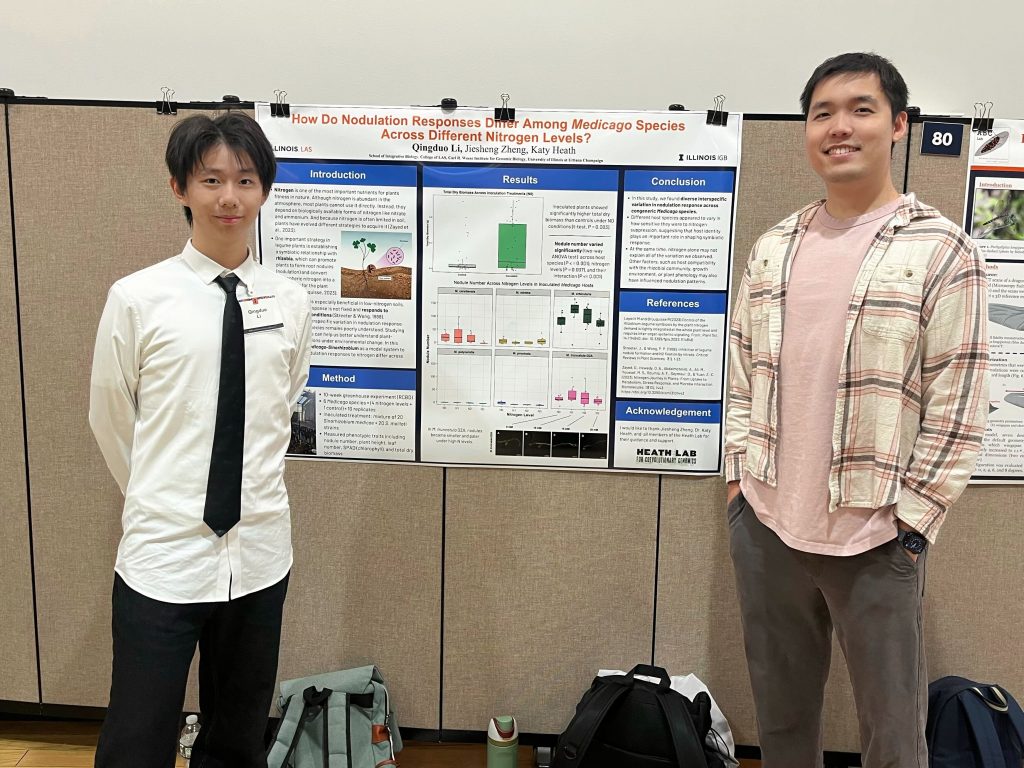
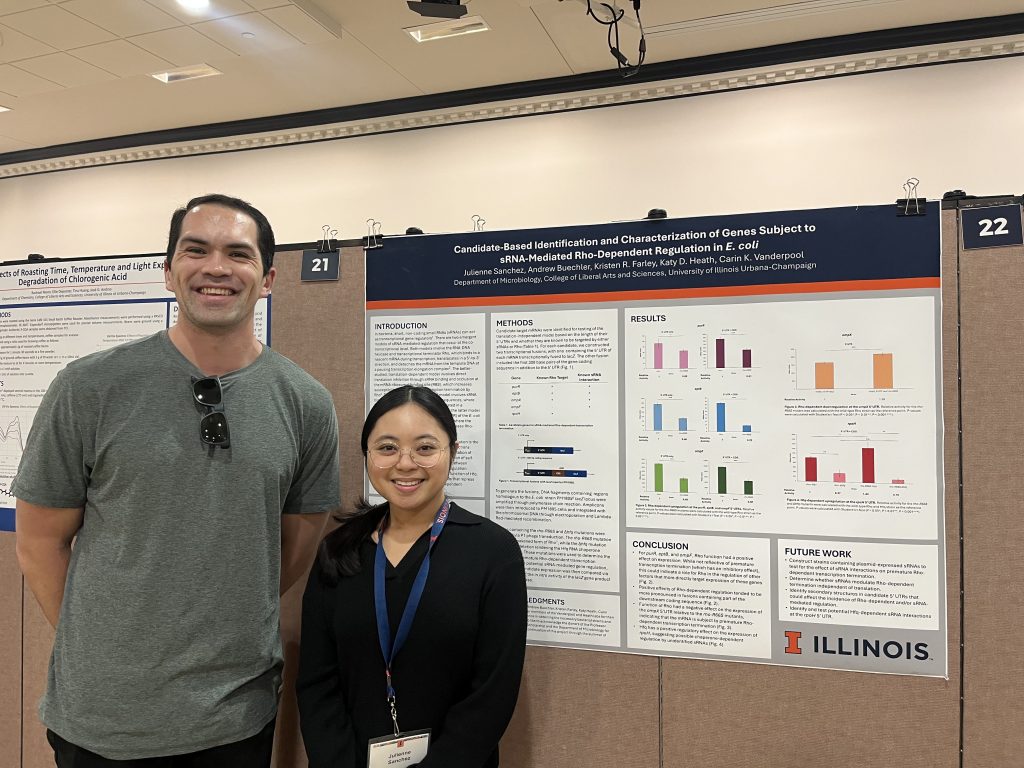


Amartya, Dave, and Julienne presented their hard work at URS and IGB. Look at those proud mentors 🙂
Natalie’s super cool project shows how genetic variation in Rhizobium partner quality can influence not only the clover hosts, but neighboring grasses and clover-grass competitive dynamics.

His first paper just came out in mBio – look for another soon on phage resistance in Rhizobium and plasmid fusion and fission. Woo!

Hats off to Connor and Allison from the Christian lab (University of Louisville) for leading this awesome review on aboveground-below ground microbial interactions!


Best of luck running and studying back in NY this year Kate – we will miss having you in the lab!


It is bittersweet that after many years of hard work, this collaborative paper (truly a co-lead situation) finally came out in PNAS right after we lost our friend, colleague, and collaborator Amy. This paper was such a labor of love, and we’re deeply thankful to have had the pleasure of working and writing with this amazing human. Love you, babe.

https://www.pnas.org/doi/epdf/10.1073/pnas.2411831122

Cole has spent the summer studying the phenotypic impacts of phage resistance on plants!