For Frank et
al’s book on Carotenoids, Advances in Photosynthesis
Research, Volume 8, Series Editor, Govindjee
Revised December 31, 1998: 11 AM version
Carotenoids in Photosynthesis: An Historical Perspective
In: "The
Photochemistry of Carotenoids", edited by H.A. Frank,
A.J. Young, G. Britton and R.J. Cogdell, published in 1999 by
Kluwer Academic Publishers, Dordrecht, pp. 1-19.
Departments of Biochemistry and Plant Biology, and Center of Biophysics & Computational Biology,
University of Illinois, Urbana, IL 61801-3707, USA
E-mail: gov@uiuc.edu;
fax: 217-244-7246
Summary
B. Carotenoids: Carotenes and Xanthophylls
C. Function: Light Harvesting and Photoprotection
II. Excitation Energy Transfer: Sensitized Fluorescence and Photosynthesis
A. Photosynthetic Yields in Different Wavelengths of Light
B. Excitation of Chl a Fluorescence by Different Wavelengths of Light
C. Resonance Excitation Transfer Theory versus Electron Exchange
Summary
This chapter presents a personal historical perspective of the
role of carotenoids in photosynthesis. It leads the readers into
the early literature on the carotenoids and photosynthesis that
are related to the discoveries on the excitation energy transfer
and to a lesser extent on photoprotection. Excitation energy
transfer from the carotenoid fucoxanthin to chlorophyll (Chl) a
was shown first in the diatoms by H. Dutton, W.M. Manning and
B.M. Duggar, in 1943, at the University of Wisconsin at Madison.
After the extensive researches of E. C. Wassink (in the
Netherlands) on this topic, the classical doctoral thesis of L.
N. M. Duysens became available in 1952, at the State University
in Utrecht. This thesis dealt with the evidence of excitation
energy transfer in many photosynthetic systems, including
anoxygenic photosynthetic bacteria. The experiments of R. Emerson
and C.M. Lewis, done at the Carnegie Institute of Washington,
Stanford, California, in the 1940s, dealt with the quantum yield
action spectra of photosynthesis. In these experiments, the
famous red drop phenomenon was discovered; further, the
authors showed here the low efficiency of carotenoids in the
photosynthesis of both green algae and blue-green algae
(cyanobacteria). In 1956, R. Stanier and his coworkers
discovered, at the University of California at Berkeley, a
special role of carotenoids in protection against death in
phototrophic bacteria. Finally, in 1962, H. Yamamoto (of Hawaii)
pioneered the role of xanthophyll cycle pigments in
photoprotection. This was followed by key experiments and
concepts from B. Demmig-Adams (1987, now in Colorado), and O.
Bjorkman (at Stanford, California), amongst others mentioned in
the text. In 1954, a 515 nm absorbance change was discovered by
Duysens (1954) and has now become a quantitative measure of the
membrane potential changes in photosynthesis. Historical aspects
of some of the basic principles of light absorption and
excitation energy transfer, and references to selected current
literature are also included in this chapter to allow the reader
to link the past with the present.
The intent in this chapter is to present a historical perspective of the two major functions of carotenoids in photosynthesis, namely light harvesting and photoprotection, with emphasis on the former. As a novice in both the history of photosynthesis and in the study of the role of carotenoids, I am unencumbered by any bias except that of personal and close associations with (1) Robert Emerson, who, with Charleton M. Lewis, measured the first most precise action spectra of photosynthesis (that included the carotenoid region) in the cyanobacterium Chroococcus (Emerson and Lewis, 1942) and the green alga Chlorella (Emerson and Lewis,1943), and discovered the enhancing effect of light absorbed by the carotenoid fucoxanthin on the quantum yield of photosynthesis sensitized by Chl a of what we now call Photosystem I (PSI) in the diatom Navicula minima (see Emerson and Rabinowitch, 1960; Govindjee and Rabinowitch, 1960; Rabinowitch, 1961); and (2) with Eugene Rabinowitch, who wrote the most detailed single-authored treatise, more than 2000 pages long, on all aspects of photosynthesis including carotenoids, published in 1945 (Vol. I), 1951 (Vol. II, part 1), and 1956 (Vol. II, part 2) (see Bannister, 1972; Brody, 1995). The personal perspective of Duysens (1989) provides an account of the discovery of the two light reactions of photosynthesis, necessary for understanding the context of the present day view of photosynthesis.
-------------------------------------------------------------------------------------------------------------
Abbreviations: Chl- chlorophyll; PSI- photosystem I; PSII- photosystem II
-------------------------------------------------------------------------------------------------------------
Retracing historical developments and comprehending
the overview of the history of the ideas are essential in
grasping the nature of scientific enquiry in any field.
From another perspective, I also believe in what Pliny,
the Younger (see a translation by Firth, 1909) implied in
Book V, Letter V to Nonius Maximus (pp. 224-225) that it
is a noble employment to rescue from oblivion those who
deserve to be remembered. In writing this chapter,
I do not even know if this would be achieved here. I
shall attempt to present a view that I consider worthy of
thought by the readers of a book that deals with the
Photochemistry of Carotenoids. I know that a great many
scientists are filled with a glow when they see others
citing and recognizing their work; they feel that what
they did was indeed useful to society.
Everything connected with color has always held, and
will continue to hold, a captivating interest for me. The
brilliant yellow pigments known as carotenes and
xanthophylls are no exception. Carotene was first
isolated in 1831 by Heinrich Wilhelm Ferdinand
Wackenroder (1798-1854). Berzelius (1837a,b) named the
yellow pigments obtained from the autumn leaves
xanthophylls (xanthos being Greek for yellow, and phyll
for leaf) as a counterpart to chlorophyll, Chl (leaf
green). Fremy (1860) reviewed the knowledge on
carotenoids at that time. By 1902, however, there were
800 publications in this field (Kohl, 1902). One of the
two major yellow pigments present in leaves was found to
be identical with the carotene from the carrot root.
Xanthophylls were discovered in algae, and one leaf
xanthophyll, lutein, was found in egg yolk (for these and
other early accounts on the carotenoids, see Lubimenko,
1927 and Smith, 1930). Strain (1938) used the name
carotenes for the hydrocarbons, and xanthophylls for
oxygenated derivatives of carotenes, but Bogert (1938)
suggested that xanthophylls be called carotenols because
of their chemical structure and because they were not
restricted to leaves. In his famous treatise on
photosynthesis, Rabinowitch (1945, 1951, and 1956)
adopted the term carotenols; thus, lutein was luteol,
violaxanthin was violaxanthol, zeaxanthin was zeaxanthol,
etc. We no longer use the ol ending, which is too
restrictive, and we are back to the terminology used by
Harold Strain.
The first separation and purification of the carotenes
and xanthophylls must be credited to the Russian botanist
Tswett (1906,1911) who invented chromatography for the
separation of the leaf pigments i.e. green Chls,
and yellow-to-orange carotenes and xanthophylls (also see
discussion of paper chromatography by Jensen and
Liaaen-Jensen, 1959). Tswett already provided the concept
of a family of many pigments, the carotenoids (carotenes
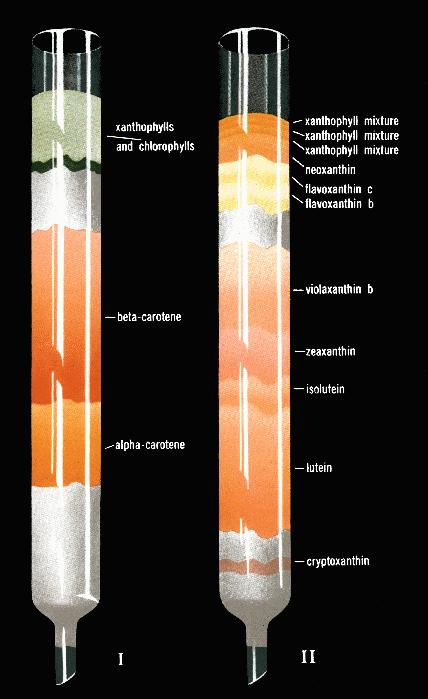
and xanthophylls). (See Figure 1,
colored, taken from Strain (1938) showing the separation
of some of the carotenoids on two systems.) This was
followed by the extensive work on the separation and
chemistry of the carotenoids by R.Wilstätter (Nobel
Prize in Chemistry in 1915, mostly for work on Chl
chemistry) (see Wilstätter and Stoll, 1913) although his
ideas on the functions of these pigments were not
substantiated. Following these early days, research on
the carotenoids was reviewed by Palmer (1922),
Zechmeister (1934, 1962), Lederer (1934), Karrer and
Jucker (1948, English translation, 1950), Goodwin (1952,
1976), Cogdell (1978; 1985: for interactions with Chls),
Britton and Goodwin (1982), Cogdell and Frank (1987),
Mimuro and Katoh (1991), Britton et al. (1995) and
Bartley and Scolnik (1995). Nobel prizes in Chemistry
were successively awarded to Paul Karrer (in 1937) and
Richard Kühn (in 1938) for their work on the structure
and chemistry of the carotenoids. A book edited by Isler
(1971) was dedicated to the memory of Paul Karrer. Kühn
(1935) showed that the carotenoids absorb in the visible
(at about 480 nm) due to the alternation of single and
double bonds, which produces a so-called Brillouin gap
when one resonance structure is dominant. It was Karrer,
however, who had recognized the symmetrical nature of the
various carotenoids (b
-carotene; lycopene; zeaxanthin) and that vitamin A was
related to half of the b
-carotene molecule (see Karrer,1934; Karrer and
Helfenstein, 1933). The nomenclature at that time was
summarized by Palmer (1934). By 1948, about 80
carotenoids were known, and structures of about half of
those were established; by 1950, total synthesis of b carotene was achieved by Karrer
and others. For chemistry of carotenes, see McKinney
(1935), Liaaen-Jensen (1978), and Packer’s two
volumes (1992a, 1992b); for antheraxanthin, see Karrer
and Oswald (1935), for spirilloxanthin, see van Niel and
Smith (1935), for xanthophylls of algae, see Strain et
al. (1944), and for carotenoids in cyanobacteria, see
Hirschberg and Chamowitz (1994). For rules on the
nomenclature of carotenoids, see IUPAC and IUB (1971,
1975).
Karrer and Jucker (1948), when dealing with the function of
carotenoids, wrote "All these investigations are still at a
preliminary stage and further researches will be required in
order to elucidate the importance of carotenoids in plants."
Similarly, Goodwin (1952) stated " with regard to formation
and function (of carotenoids), knowledge is rudimentary".
However, by this time Dutton and Manning (1941) in Wisconsin had
already shown that light energy absorbed by fucoxanthin was used
efficiently for photosynthesis in the diatom Nitzschia
closterium, and Dutton et al. (1943) had clearly established
that this process took place by transfer of energy absorbed by
fucoxanthin to Chl a, because excitation of fucoxanthin
led to Chl a fluorescence (i.e., the phenomenon of
sensitized fluorescence was observed). This was a clear case of
the light-harvesting function of one of the carotenoids in
vivo (also see Dutton, 1997). Further coverage of the history
on this topic will be presented in section II of this chapter.
Carotenoids are known to have another major function, i.e. that of photoprotection of reaction centers, pigment-protein antennae, and cells and tissues (see Krinsky, 1968, 1979). The work of Roger Stanier and his coworkers provided the most compelling evidence for the belief that carotenoids perform a photoprotective function. In 1955, Griffiths et al. discovered that a blue-green (BG) mutant of the non-sulphur purple bacterium Rhodopseudomonas (now Rhodobacter) sphaeroides, which is deficient in colored carotenoids, is photosensitive in the presence of air. The mutation was lethal. It was suggested that carotenoids are universally associated with photosynthetic systems because they protect these systems against photodynamic damage catalyzed by bacterioChl (in photosynthetic bacteria) or by Chl (in plants and algae). More recently, a third function was discovered: in higher plants exposed to strong light. Zeaxanthin and antheraxanthin, formed from violaxanthin by the xanthophyll cycle (Yamamoto et al., 1962) increase non-radiative dissipation of energy as heat in the pigment bed of the antenna of Photosystem II (PSII) (Demmig et al., 1987; Gilmore et al., 1995). Further discussion on this topic will be presented in section IV of this chapter (also see Horton et al., 1994, Demmig-Adams et al., 1996; Yamamoto and Bassi, 1996; Gilmore, 1997).
The reader is referred to Sieferman-Harms (1987a,b) for
earlier discussions on the light harvesting and photoprotective
functions of carotenoids.
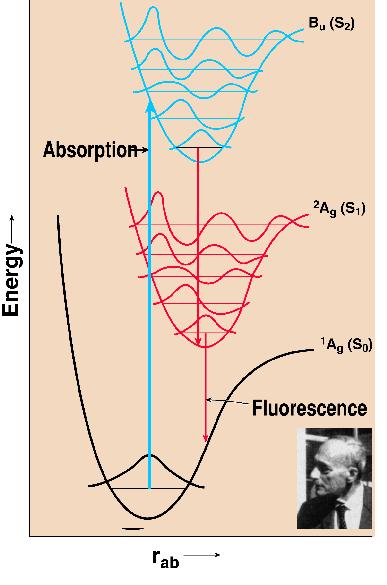
Pigment excitation occurs after the absorption of light. This promotes the molecule from the ground state (So) to an excited state (Sn). All further reactions occur after de-excitation of this higher excited state. This upward transition occurs in 1-2 fs in visible light. A historical suggestion was that of James Franck (1925, who had shared, with Hertz, the 1925 Nobel Prize in Physics for the experimental verification of the quantum theory). He argued simply that because of the large masses of the nuclei in a molecule, their relative momentum cannot be directly affected by an electronic transition, so that those transitions will be most likely that conform most closely to a Principle. The nuclei do not move during an electronic transition. Thus, on a diagram of energy (ordinate) versus distance between the nucleii of a diatomic molecule (abscissa), this transition is vertical promoting the electron from the lowest vibrational state of a molecule in the ground state to a higher vibrational state of the excited state (e.g. S1 or S2) of the molecule (Figure 2). The molecule in the excited state then dissipates immediately (within 10 to 100 fs) some energy as heat and the electron reaches the lowest vibrational level of the excited state. When the molecule relaxes to the ground state giving off light (fluorescence), it generally occurs at longer wavelength than the absorbing wavelength (Rotverschiebung, the red shift, Franck, 1927). The Franck-Condon principle, then, explains the observed red shift (Stokes, 1852) of the fluorescence spectrum from the absorption spectrum. The history of how the principle became known as the Franck-Condon principle was beautifully presented by Condon (1947). The original idea is in a paper at a Faraday Society meeting in London by Franck (1925); the proofs of this paper were sent to his student Hertha Sponer, who was then at the University of California at Berkeley on an International Education Board Fellowship. She generously shared the proofs with Condon; he was able to generalize Franck’s ideas (Condon, 1926). Condon (1947) states:"This work was all done in a few days. Doctor Sponer showed me Franck’s paper one afternoon, and a week later all the quantitative work for my 1926 paper was done." With carotenoids, one does not usually observe by conventional absorption spectroscopy, transitions involving the first singlet excited state, (S1) but mainly the second singlet state (Bu or S2). The transition from the So state to S1 is optically forbidden (for a fuller discussion, see Chapter 8, this volume; also see a review by Koyama, 1991). Fluorescence of carotenoids in general is very weak (S1 to S0 transition).
II. Excitation Energy Transfer: Sensitized Fluorescence and
Photosynthesis
The first major function of carotenoids is to act as an
accessory pigment, i.e., to capture light and transfer the
energy to Chl a to drive photochemistry. The methods used
to obtain evidence for this are basically two: (1) measurement of
action spectrum of photosynthesis in the region carotenoids and
Chls absorb and evaluation of the quantum efficiency of light
absorbed by carotenoids in photosynthesis; and (2) measurement of
action spectrum of Chl a fluorescence in the region where
carotenoids and Chls absorb and evaluation of the quantum
efficiency of excitation energy transfer from the carotenoids to
Chl a. The latter technique is called the sensitized
fluorescence method. If the energy donor is fluorescent, one
would observe decreases (quenching) in donor fluorescence and
increases in acceptor fluorescence when the donor is excited,
whereas excitation of the acceptor would lead to acceptor
fluorescence only; this method was first applied by Cario and
Franck (1923) in gases. Since then it has been successfully used
in liquids, solids, proteins and photosynthetic systems (see e.g.,
Knox, 1975; Stryer, 1978; Pearlstein, 1982; van Grondelle and
Amesz, 1986; Frank et al., 1991; van Grondelle et al., 1994).
A.Photosynthetic Yields in Different Wavelength Regions
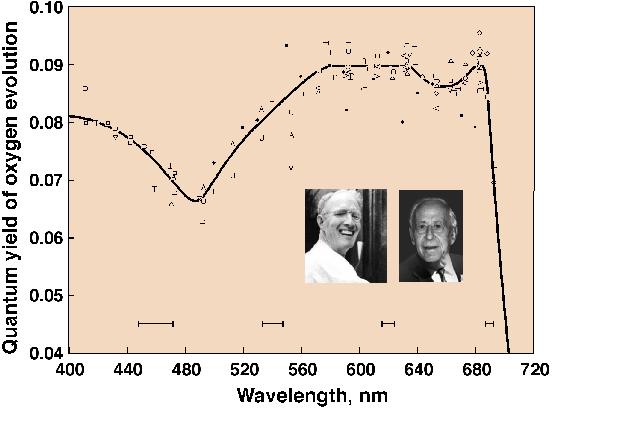
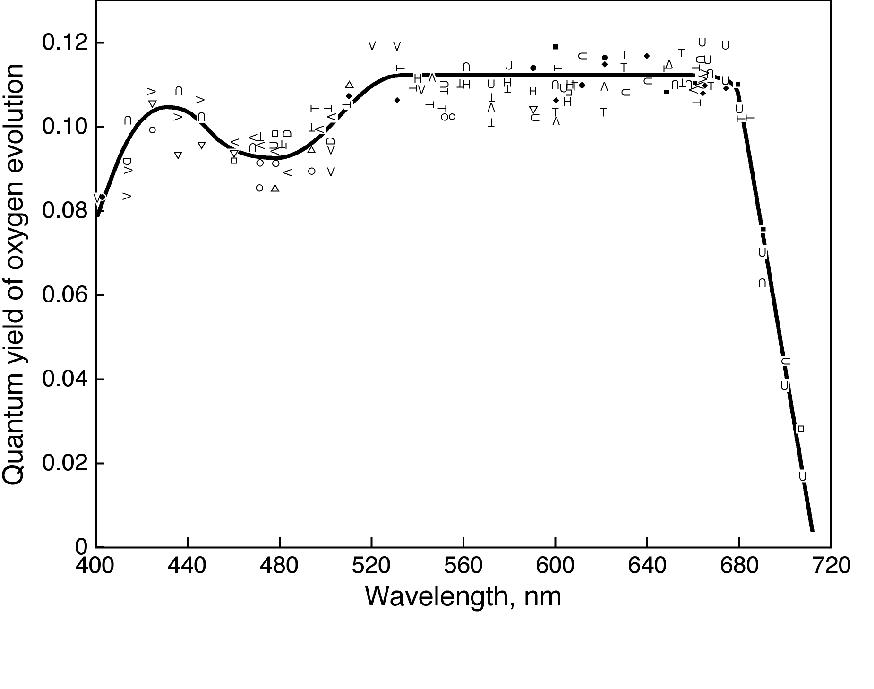
Engelmann (1883, 1884) was an ingenious scientist (see Kamen, 1986). He projected the visible spectrum on to green, red and brown algae, mounted on the stage of a microscope, and used the number of aerotactic motile bacteria accumulating in the different wavelengths of the light as an indication of the rate of oxygen evolution. He concluded that light absorbed by various accessory pigments (including carotenoids, particularly fucoxanthin) was used for photosynthesis. Warburg and Negelein (1923), using precise manometric methods, measured absolute quantum yields of oxygen evolution by the green alga Chlorella in different colors of light. Although the absolute quantum yield of oxygen evolution in blue light, where both carotenoids and Chls absorb light, was later questioned by others, it was slightly lower than in the red where only Chls absorb light. Thus, this experiment indicated that, although carotenoids contribute to photosynthesis, their efficiency is somewhat lower than that of Chls. It was Montfort (1936, 1940) who compared, although rather crudely and from unreliable experiments, absorption by various extracted photosynthetic pigments and oxygen evolution in various colors of light, and concluded that light absorbed by fucoxanthin of marine brown algae is fully utilized in photosynthesis. The first extensive and reliable measurements on the quantum yield of oxygen evolution as a function of wavelength of light (i.e., of the action spectrum of photosynthesis) were, however, carried out by Emerson and Lewis (1942, 1943) on the cyanobacterium Chroococcus and the green alga Chlorella (see Figure 3). These experiments were done with a large home-built monochromator with the grating obtained from Mt. Wilson observatory and the use of the most precise manometry, where 0.01 mm pressure changes, due to oxygen evolution, could be measured! Independent of Emerson’s work, Dutton and Manning (1941) carried out similar experiments with the diatom Nitzschia closterium (now Phaeodactylum tricornutum), using a dropping mercury electrode. Their conclusions were that fucoxanthin in the diatom is almost 90% efficient, whereas the carotenoids are about 40-50% efficient in Chlorella, and much less efficient (perhaps, only 10%) in Chroococcus. Although Wassink and Kersten (1945,1946) came to the same conclusion as Dutton and Manning (1941), it was Tanada (1951), a student of Emerson, who provided the most thorough and precise data. He used the Emerson-Lewis monochromator, and showed a very high quantum yield of oxygen evolution at 500 nm, where fucoxanthin absorbs most of the light (see Fig. 4). On the other hand, Haxo and Blinks (1950) made a large number of action spectra of photosynthesis plotted per incident photons in many marine algae, but were able to make only qualitatitive statements regarding energy transfer from carotenoids to Chl a. They concluded that carotenoids were relatively inactive in the green alga Ulva, but considered that some carotenoids must be active in photosynthesis in some systems. For a review on the action spectra of photosynthesis, see Fork and Amesz (1969).
Measurements on the action spectra of photosynthesis do not
distinguish between the direct photochemistry by the carotenoids
versus that by Chl a after excitation energy transfer from
them to the Chls. This distinction is possible only from
measurements on excitation energy transfer.
B. Excitation of Chl a Fluorescence by Different Wavelengths of Light: Sensitized Fluorescence
Vermeulen et al. (1937), in the laboratories of L. S. Ornstein
of Utecht and A. J. Kluyver of Delft, published their results on
the intensity of fluorescence per quantum absorbed as a function
of wavelength of light for the green alga Chlorella.
Although the measured quantum yields of Chl a fluorescence
were too low to be true, the yield of Chl a fluorescence
after excitation by 496 nm light was about 20% lower than after
excitation at 607 nm. The authors stated that both the quantum
yield of the Chl a fluorescence and that of photosynthesis
(Warburg and Negelein, 1923) was independent of wavelength in the
region where only Chl a absorbs. Since the data show
10-30% decreases in the blue (see Vermeulen et al, Table III), we
can conclude that carotenoids did not transfer 100% of their
excitation energy to Chl a although the authors did not
make any comment on this problem.
The first and clear evidence for excitation energy transfer
from fucoxanthin leading to Chl a fluorescence was
obtained by Dutton et al. (1943). It was demonstrated that light
absorbed by fucoxanthin was almost 90% as efficient in producing
Chl a fluorescence as was light absorbed by Chl a
itself. This was the clearest pioneering paper dealing with
sensitized fluorescence evidence for excitation energy transfer
in photosynthesis (see Dutton, 1997, for the experimental
background prior to the actual experiment). Excitation energy
transfer, in general, in photosynthesis was implied already in
the paper of Gaffron and Wohl (1936) when they were explaining
the photosynthetic unit experiments of Emerson and Arnold
(1932a,b). Further, Oppenheimer (1941) had called it internal
conversion while thinking about the still earlier unpublished
experiments of William Arnold (see Knox, 1996, for the history of
this work, as well as Arnold and Oppenheimer, 1950). Wassink and
Kersten (1946) confirmed the conclusion of Dutton et al. (1943)
on excitation energy transfer from fucoxanthin to Chl a.
Although Van Norman et al. (1948) did not really discuss
excitation energy transfer from phycoerythrin (a phycobilin, not
a carotenoid) to Chl a in the red algae they had examined,
it was clear that the higher yield of red fluorescence by
excitation with green light, absorbed by phycoerythrin, than by
red light, absorbed by Chl a, suggested efficient
excitation energy transfer from phycoerythrin to Chl a.
Excellent evidence for this transfer was published by French and
Young (1952), and was known to and fully recognized by L. N. M.
Duysens (1951, 1952). No discussion of energy transfer from
carotenoids to Chl a is available in the papers of French
and coworkers. The classical work of Duysens (1951, 1952)
established that: (1) carotenoids transfer 35-40% of their
absorbed energy to bacterioChl a in the B890 complex of Chromatium
strain D, and about 50% to bacterioChl a in the B890
complex of Rhodospirillum molischianum; (2) about 40% of
energy absorbed by carotenoids is transferred to Chl a in
green algae; and (3) about 70% of energy absorbed by fucoxanthin
is transferred to Chl a in diatoms and brown algae.
In none of the early experiments, except for the work on
fucoxanthin, was any distinction made between carotenes and
xanthophylls. In 1956, Arnold and Meek presented their work on
the depolarization of Chl a fluorescence (see Perrin,
1926, 1929), thus supporting clearly the concept of excitation
energy migration in photosynthesis. To me, this was an important
experiment of its time.
When I joined the research group of Robert Emerson in 1956,
Emerson was very keen that I work on the problem of the separate
roles of carotenes and xanthophylls in photosynthesis. I grew
several types of algae (Tribonema; Muriella; Tolypothrix)
in different colors and intensities of light and extracted
carotenes and xanthophylls and monitored the variations in the
ratios of the two groups of carotenoids under various
experimental conditions. Unfortunately, for me, Emerson was not
interested in measuring action spectra of Chl a
fluorescence, but was only interested in measuring quantum yield
action spectra of photosynthesis, an art he had perfected. The
progress on my research was extremely slow due to my impatience,
the tedious nature of manometry and difficulties in measuring
absolute quantum yield of photosynthesis in low intensities of
different wavelengths of light. My work was never finished in
spite of piles and piles of data I had collected. (I moved on to
other research after Emerson’s death on February 4, 1959.)
However, Goedheer (1969a,b; see his review, 1972), also from the
same laboratory as Duysens, published a paper on chloroplasts
treated with petroleum ether (this treatment selectively removes
carotenes, not xanthophylls) and concluded that, in red algae and
in cyanobacteria whose phycobilins he had also removed, b carotene transfers energy to Chl a
(of PSI with 100% efficiency, whereas the b
carotene of green algae and greening leaves transfers energy to
Chl a of both PSI I and II. Surprisingly, Goedheer
concluded that xanthophylls in these organisms do not transfer
any energy to Chl a. The observed peaks for the
carotenoids were at 471 nm and 504 (or 506) nm, at 77K in the
action spectra of Chl a fluorescence. I believe there is a
need for further research on this topic in intact systems without
such solvent treatments, as used by Goedheer. There is, however,
evidence that lutein, a xanthophyll, transfers energy to Chl a
with 100% efficiency in the isolated light harvesting complex
(LHC) of Lactuca sativa (see Siefermann-Harms and
Ninnemann, 1982). Although many authors state that violaxanthin
transfers excitation energy to Chl a, Barrett and Anderson
(1980) could not detect any significant excitation energy
transfer in the green Chl a/c-violaxanthin protein from
the brown alga Acrocarpia paniculata.
After I finished my PhD under Eugene Rabinowitch and had
established that Chl a was in both the photosystems
(Govindjee and Rabinowitch, 1960) and that Chl a
fluorescence measurements can be used to support the existence of
two light reactions and two photosystems (Govindjee et al., 1960,
Duysens, 1989), I went back to measure the action spectra of Chl a
fluorescence, but not so much from the point of deciding the role
of carotenoids in photosynthesis, but of simply using Chl a
fluorescence as an intrinsic, sensitive, and non-invasive probe
of photosynthesis (see Govindjee, 1995). A major interest was in
using temperature dependence of fluorescence down to liquid
nitrogen and helium (4K) temperatures. Figure
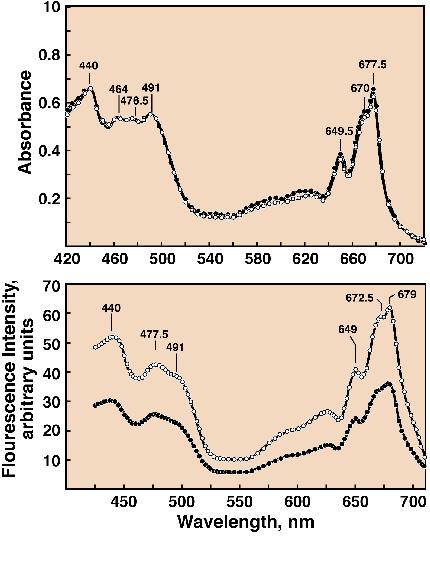
5 (Cho and Govindjee, 1970a) shows the
absorption spectra and action spectra of Chl a
fluorescence in the green alga Chlorella pyrenoidosa at 77
and 4K. Several bands can be observed including the one at 491 nm
from carotenoids. Excitation energy transfer from carotenoids to
Chl a is clearly indicated, but no further information is
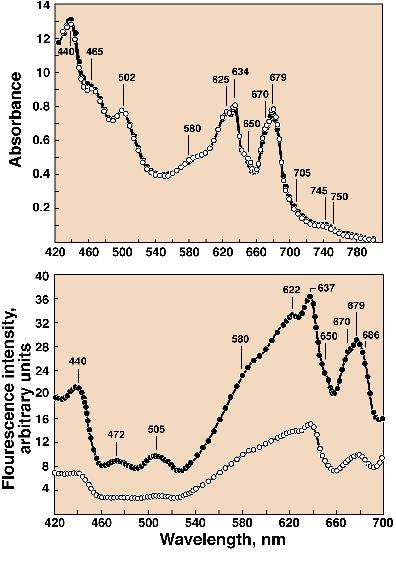
available. With the blue-green alga (cyanobacterium) Anacystis
nidulans, bands at 472 nm and 505 nm, due to carotenoids, are
observed (Figure 6, Cho and
Govindjee, 1970b; also see Kramer et al., 1981, for similar data
on spinach and barley) in the action spectra of Chl a
fluorescence, again showing energy transfer from carotenoids to
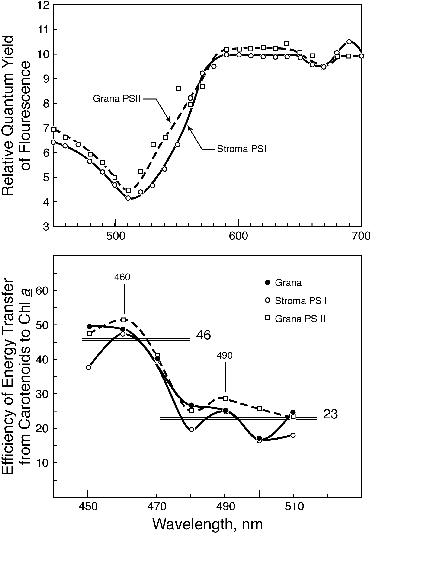
Chl a. In grana and stroma lamellae fractions from
thylakoids, Gasanov et al. (1979) calculated the efficiency of
excitation energy transfer from the carotenoids (without
distinction between carotenes and xanthophylls) to Chl a (Figure 7). It appears that there are two
pools of carotenoids, one absorbing at shorter wavelengths and
transferring energy to Chl a with an efficiency of about
40-50% and another at slightly longer wavelengths transferring
energy, with an efficiency of about 20-25%. This conclusion has
never been confirmed or pursued, and the question of the precise
roles of carotenes and xanthophylls in light harvesting remains
still an open question worthy of research.
It was already known in the nineteenth century that a part of
another carotenoid (peridinin) is bound to a protein in vivo
in dinoflagellates (Schuett, 1890). The excitation spectrum of
Chl a fluorescence in the chromoprotein from Amphidinium
carterae, obtained by Haxo et al. (1976), showed that light
absorbed by peridinin is transferred efficiently to Chl a.
Song et al. (1976) showed 100% efficiency of energy transfer from
peridinin to Chl a in Chl a-proteins of two
dinoflagellates, a Glenodinium sp. and Gonyaulax
polyedra. Further, efficient energy transfer has been
reported from siphonaxanthin (absorbing in the green region) to
Chl a in the thalli of the green algae Ulva japonica
and Ulva pertusa (Kageyama et al., 1977) and in the
isolated green protein, containing siphonaxanthin, from Codium
(Anderson, 1983; also see a review by Govindjee and Satoh,
1986).
C. Resonance Excitation
Transfer Model Compared to Electron Exchange
As already mentioned, evidence for excitation energy transfer
to Chl a was established, first from fucoxanthin and then
from peridinin, as well as from b
-carotene. This raises question of how excitation energy moves
from one molecule to another. In 1940, William Arnold had
observed excitation energy transfer from phycocyanin to Chl a
and had discussed it with JR Oppenheimer (see Arnold, 1991, p.
77). Knox (1996) has traced the history of Arnold’s
contribution; he states that the R-6 dependence of
excitation energy transfer (to be evolved later by Förster) must
have been evident to Oppenheimer in whose 1941 paper the
conclusion of Arnold and Oppenheimer (1950) was already stated.
There are two major theories for exciton transfer: (A) Förster’s
resonance energy transfer mechanism (Förster,
1946,1948, 1965) (called the Heller-Marcus mechanism in the
crystal field) that depends upon the transfer of excitons where
the decay of the excited state in the donor molecule is coupled
with the upward transition promoting the ground state of the
acceptor to the excited state. Energy (i.e., hole and
electron together) is transferred from one molecule to the other.
This mechanism is based upon a dipole -(induced) dipole
interaction and includes the following: (1) an R-6
dependence of energy transfer, where R is the distance between
the donor and the acceptor; (2) an appropriate orientation of the
dipoles for a efficient transfer; and (3) a good proximity of the
energy levels, as measured usually by the overlap integral of the
absorption spectrum of the acceptor molecule and the fluorescence
spectrum of the donor molecule (see e.g., Knox, 1975; van
Grondelle and Amesz, 1986). [The R-6 dependence was
clearly proven in in vitro by studying excitation energy
transfer from an a - naphthyl group at
the carboxyl end of a polypeptide to the energy acceptor dansyl
group at the amino end, when the distance was changed by spacers
of oligomers of poly-L-proline (Stryer and Haugland, 1967).] (B) Dexter’s electron exchange
mechanism (Dexter, 1953); here, there is electron
exchange, i.e., the movement of an electron from the LUMO
(lowest unoccupied molecular orbital in the ground state) in the
donor molecule to the vacant LUMO in the acceptor simultaneous
with the movement of the electron from the HOMO (highest occupied
molecular orbital) of the acceptor molecule to the vacant HOMO of
the donor molecule (called the Wannier mechanism in the crystal
field). This mechanism requires extremely close proximity of the
donor and acceptor molecules. The energy level diagram (the
Jablonski diagram, see Jablonski, 1935) shows that the energy
level of the optically allowed Bu (S2)
state of the carotenoid molecule is higher than the S2
level of Chl or BChl. In general, however, the lifetime of the S2
state of the molecules is too short and most of the
de-excitation occurs by loss of heat and the attainment of the S1
state. There are, however, reports that excitation energy
transfer may be possible originating from S2 (see e.
g., Chapter 8, this volume). The consensus is that the lower
level of carotenoids is expected to be involved in excitation
energy transfer. Now, since the 2Ag (S1) to
Ag (S0) is optically forbidden, it has been suggested
that the singlet energy transfer reaction, involving the S1
states, must take place through the electron exchange mechanism.
For a review see Frank and Cogdell (1993).
I consider it interesting to mention an old idea of Platt
(1959) who predicted that energy can be transferred from
carotenoids to Chls, but entertained the possibility of energy
transfer from Chl to a charge-separated state involving carotene.
He had predicted large red shifts from an absorption at 480 nm,
to the orange-red region, and then to 1100 nm for various states
of carotenes. No one since then has provided any specific
experimental support for Platt’s ideas.
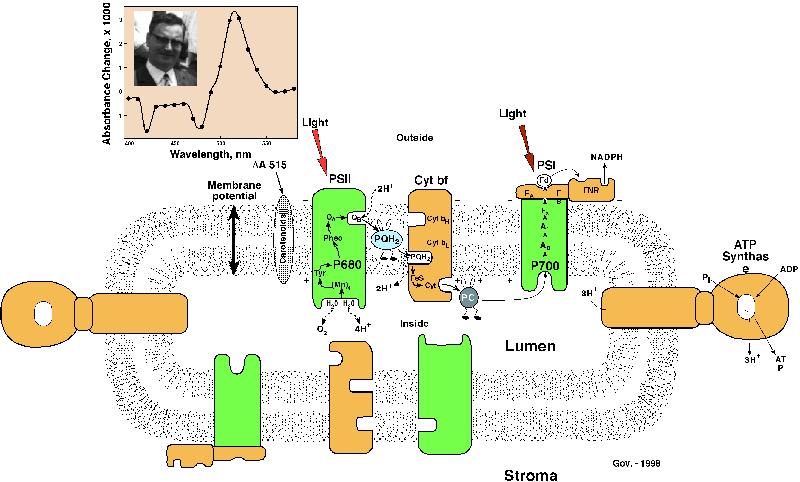
III.The 515 nm Effect: Carotenoids as a
Microvoltmeter
An outline of the history of the role of carotenoids in
photosynthesis would certainly be incomplete without a discussion
of the so-called 515 nm effect (D A
518 as some call it). When a pigment is placed in an electric
field, its absorption spectrum is shifted because the field
changes the energy levels of the pigment. This is so-called Stark
effect. During photosynthesis, electrons are transferred from one
side of the thylakoid membrane to the other side since the
primary electron donors (P680, P700, P870, etc) are located on
one side and the stable electron acceptors on the other side (Figure 8). This produces a membrane
potential (electric field). Thus, the light-absorbing properties
of the pigments present in the membrane are then affected as a
result of the Stark effect; this produces what we call electrochromism.
Carotenoids are affected in this way and are responsible for a
major portion of the absorption change (D
A 515 or D A518).
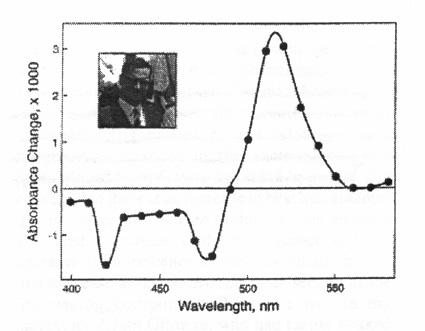
A positive absorbance change around 515 nm was discovered in
the green alga Chlorella, in a leaf, in the thalllus of a
marine alga, and in the blades of Valisneria. (Figure 9) by Duysens (1954) when
he was a visiting fellow at the Carnegie Institute of Washington
at Stanford, after his brief stint as a fellow at the UIUC,
Urbana, Illinois, with Eugene Rabinowitch. This positive
change was accompanied by negative changes at 480 nm and 420 nm;
the latter was assigned to cytochrome f. It was Strehler (1957)
who suggested its relationship to carotenoids (also see Govindjee
and Govindjee, 1965). Wolff et al. (1969), in the laboratory of
H.T. Witt, pioneered the relationship of the 515 nm change to the
fast charge separation processes at the reaction centers because
they observed that the change occurred within nanoseconds after a
flash of light. It was later shown by HT Witt and coworkers that
about 50% of the fast change arises from PSI and the other 50%
from PSII (see e.g., a review by Witt, 1975). According to
the chemiosmotic hypothesis of Peter Mitchell (1961), proton
motive force (i.e., D pH and D Y ) is used to
produce free ATP. The 515 nm change was found to decay faster in
the presence of gramicidin D, an uncoupler of
photophosphorylation, as expected if the change is a monitor of
membrane potential (Junge and Witt, 1968). I note that in this
paper, the authors assumed that they were monitoring changes only
in Chl b at 515 nm. Jackson and Crofts (1969) made another
important observation in bacteria. They found a shift in the
carotenoid spectrum (523 minus 509 nm absorbance change) in
darkness when a potential is generated that is positive with
respect to the inside of the chromatophores; the shift mimicked
that observed as a response to light. De Grooth et al. (1979)
observed a flash number dependency of the biphasic decay of the
electrochromic shift of the carotenoids, related to the changes
in the membrane potential, in the chromatophores of Rhodobacter
sphaeroides. The carotenoid absorbance change is accepted now
to be a monitor of the membrane potential.
The hypotheses to explain how carotenoids play a role in
protecting plants against damage by excess light have been
discussed very extensively. Crucial work on the topic of
photoprotection was done in the summer of 1954 in C. B. van
Niel’s Lab at the Hopkins Marine Station by Roger Stanier
and his colleagues (see Griffiths et al.,1955; Sistrom et al.,
1956; Stanier, 1960; the participation of Germaine
Cohen-Bazire in this work was acknowledged). Based on experiments
with a blue-green mutant of Rhodopseudomonas (now
Rhodobacter) sphaeroides that contained no colored
carotenoids, it was suggested that "the primary function of
carotenoid pigments in phototrophs is to act as chemical buffers
against photooxidation of other cell constituents by (B)Chl, thus
conferring a high degree of immunity to endogenous
photosensitization." The mutant was unable to live normally.
The mechanism of action was shown later to involve removal of
singlet oxygen by carotenoids, and the formation of triplet
states of carotenes (see reviews by Krinsky, 1968, 1971). We
shall not discuss this function further. However, we note that
Mimuro et al. (1995) have detected two b
-carotene molecules in the reaction center of PSII by
fluorescence and linear dichroism spectroscopy. It was shown that
the two b -carotene molecules in the
reaction center of PSII are spectrally different and transfer
excitation energy to Chl a at 77K; one has an absorption
band at 489 nm, and the other has bands at 506 and 467 nm. It is
assumed that these b -carotene
molecules must function to protect the reaction center Chls from
damage. (see e.g., Telfer et al., 1994). Further, recently,
Trebst and Depka (1997) have shown that b -carotene is essential for the assembly of
the D1 protein into functional PSII.
One of the several mechanisms by which plants protect
themselves against excess light is by dissipating excess energy
as heat through the participation of the xanthophyll cycle and
the pH gradient (see reviews by Horton et al., 1994, 1996;
Demmig-Adams et al., 1996; Demmig-Adams and Adams, 1996; Gilmore,
1997; Gilmore and Govindjee, 1999, and Chapters 14 and 15, this
volume). De-excitation of a molecule, excited by light, occurs by
fluorescence, heat, excitation energy transfer, or
photochemistry. At high light intensities, when photosynthesis is
saturated, unusual photochemistry can take place that can lead to
damage of the photosynthetic apparatus. This could be avoided if
there were a mechanism to increase energy loss as heat or
fluorescence. However, during exposure of plants to high light,
Chl a fluorescence intensity has been shown to decrease
(fluorescence quenching). One of the current suggestions is that
excess light somehow promotes the formation of zeaxanthin from
violaxanthin, with antheraxanthin as an intermediate. It is now
generally believed that it is mostly zeaxanthin (or
antheraxanthin) that removes the excess energy from the excited
Chls and loses this energy as heat. The history of the
xanthophyll cycle goes back to Sapozhnikov et al. (1957), who
first observed that violaxanthin levels changed in light/dark or
high light/low light treatments in Sakhlin buckwheat (Polygonum
sacchalinense F. Schmidt), in cyclamen (Cyclamen persicum
L), broad bean (Vicia faba L.) and medicinal dandelion (Taraxacum
officinale L s.l). They thought that violaxanthin was
converted into lutein and speculated on the possibility that this
may have significance for oxygen evolution in photosynthesis.
Yamamoto got involved at this stage and it was he who discovered
the currently accepted stepwise and cyclical pathway, now known
as the xanthophyll cycle (Yamamoto et al., 1962; also see
Yamamoto, 1979). The stepwise pathway excluded the possibility of
involvement in photosynthetic oxygen evolution. Further, the
kinetics was too slow and Yamamoto showed soon thereafter that
the effect of light was indirect. There have been many other
researchers in this field that are also currently active. It was
Barbara Demmig, Olle Björkman and their coworkers (see
Björkman,1987; in this paper the author mentions two unpublished
manuscripts of Demmig and Björkman; Demmig et al., 1987: work
done in the pharmaceutical laboratory of Professor Czygan;
Björkman and Demmig-Adams, 1994) who suggested that the excess
energy is lost as heat in the antenna complexes, and related the
phenomenon to the xanthophyll cycle. Barbara.Demmig-Adams (in a
recent personal note to me) also credits Professor
Lichtenthaler's earlier papers relating Chl a fluorescence
changes to the xanthophyll cycle. The concept that there is an
increase in heat loss assumes that the observed decrease in
fluorescence intensity is indeed a decrease in the fluorescence
yield. A decrease in fluorescence intensity could also be due to
a decrease in the absorption cross-section of the fluorescing
component. While he was in my laboratory Adam Gilmore, who had
earlier worked with Harry Yamamoto and Olle Björkman, made the
measurements on the lifetime of Chl a fluorescence that
directly measures the quantum yield of fluorescence (Gilmore et
al., 1995, 1996,1998). We established that in thylakoid samples
there is a dimmer switch in which the fraction of a long-lifetime
component (a 2 ns component) of fluorescence decreases with a
concomittant increase in the fraction of a short-lifetime
fluorescence component (a 0.4 ns component). The latter component
was suggested to have increased dissipation of energy as heat
because, in these experiments, photochemistry was blocked by the
use of a herbicide, diuron. It is also now clear that both a
proton gradient (or a low internal pH) and the presence of
zeaxanthin (or antheraxanthin) are required for this process. The
protons are suggested to have a dual role: (i) activation of
de-epoxidase that leads to increased conversion of violaxanthin
into zeaxanthin, and (ii) conformational changes that lead to
efficient binding of zeaxanthin or (antheraxanthin) on antenna
complexes where dissipation of energy as heat takes place (Figure
10). Whether the
heat loss occurs via zeaxanthin directly, as stated above, or is
induced in Chl by association with zeaxanthin remains an open
question. The possibility that S1 state of zeaxanthin
(that has not yet been directly observed) lies below the S1
state of Chl a (Frank et al. 1994; Owens, 1996) is a grand
and a reasonable hypothesis. It makes it easy to accept that Chl a
can transfer excitation energy to zeaxanthin. Several
investigators have now established that the photoprotection
mechanism need not require light-harvesting complex IIb (see e.g.,
Gilmore et al., 1996; Briantais et al., 1996). Thus, it was
suggested that the inner antenna complexes are involved. Bassi et
al. (1993) have shown that the xanthophyll-cycle pigments are
preferentially associated with the inner antenna Chl a
complexes (CP) 26 and 28. This idea was elegantly supported by
Crofts and Yerkes (1994) when they compared the amino acid
sequences of the various light harvesting complexes (LHCIIb,
CP26, CP28, etc). Current research on the mutants of Chlamydomonas
reinhardtii and Arabidopsis thaliana, that are blocked
in the interconversions of the xanthophyll-cycle pigments, are
providing information on the molecular mechanism of the
photoprotection process (see e.g., Niyogi et al., 1997a,b,
1998; Pogson et al., 1996, 1998). A possibility has been
raised that lutein may also be important in the mechanism of
photoprotection in Chlamydomonas, but not in Arabidopsis.
What is certain is that both b -carotene and the xanthophyll fucoxanthin transfer excitation energy to Chl a; b -Carotene, in addition, protects against photochemical damage of the reaction centers, and the xanthophyll zeaxanthin protects plants against excess light by initiating reactions, in combination with those initiated by pH gradient, that lead to loss of excess energy as heat. Much research is needed to prove the roles of other carotenoids (e.g., lutein, violaxanthin, and others). It is however currently assumed that violaxanthin acts as a light harvester, i.e., transfers energy to Chl a, and that lutein may indeed substitute for zeaxanthin in some systems. Research on both the mechanism of excitation energy transfer from Chls to carotenoids and vice versa is ongoing. The availability of structures at atomic levels is certainly important for this purpose. For example, the atomic level structure of the reaction center of photosynthetic bacteria shows a carotenoid (1,2-dihydroneurosporene). (Deisenhofer and Michel, 1989). In addition, the peridinin-Chl a complex from Amphidinium carterae also shows precisely where the carotenoid peridinin is located in this antenna complex (Hofmann et al., 1996; see the colored photograph, Figure 11). Kühlbrandt et al. (1994) have provided the atomic level structure of LHCIIb, the major light harvesting Chla/Chlb complex of plants and green algae,; this has allowed the rationalization of the proposed mechanisms of excitation energy transfer among the Chls.
In my laboratory, Xiong et al. (1996, 1998) have produced a
hypothetical structural model of PSII reaction center where the
two b -carotenes molecules are
parallel to each other, although there is reason to believe that
they may be perpendicular to each other (see e.g., Mimuro
et al., 1995; Figure 12). The entire
field seems to be still in its infancy and the present book
should help encourage further research to unfold the relationship
between the molecular structure and the molecular function of all
the carotenoids.
Drs. Harry Yamamoto and Barbara Demmig-Adam have provided
important information on section IV of this chapter. However, all
omissions and errors are mine. I am thankful to all of my current
photosynthesis colleagues at the University of Illinois at Urbana
for social and intellectual support. In particular, I owe deep
gratitude to Drs. John Whitmarsh, Colin Wraight, Don Ort and Tony
Crofts for being there for intellectual interactions.
Photosynthesis training of graduate students and post-doctoral
associates at Urbana was recently supported by NSF DBI 96-02240.
Figure Legends
Figure 1. Separation of
leaf carotenes and leaf xanthophylls by chromatographic
adsorption, obtained in 1938 by Harold Strain is illustrated
above. I. Separation of a
carotene from b carotene by adsorption
of petroleum ether extracts of leaves on a magnesia column. II.
Separation of leaf xanthophylls by adsorption of a dichloroethane
solution of these pigments on a magnesium column; note the
separation of violaxanthin, zeaxanthin and lutein, mentioned in
the text. The figure is taken from Strain (1938, p. ii,
frontpiece).
Figure 2. Franck Condon
principle as applied to carotenoids. The photograph of James
Franck, the co-discoverer of the principle is shown as an inset,
and was taken by the author in 1963 when Franck attended the
Airlie House Conference on "Photosynthetic Mechanisms of
Green Plants", organized by Bessel Kok and Andre Jagendorf.
Figure 3. The first
detailed quantum yield action spectra of oxygen evolution in the
green alga Chlorella pyrenoidosa, obtained by Emerson and
Lewis (1943) using Warburg manometry. Nineteen sets of
experiments were made and the phenomenon of the red drop
(drop in the quantum yield of oxygen evolution, beyond 680 nm)
was discovered; this later led to the discovery of the two
pigment system and two light reaction scheme when Emerson and
co-workers discovered the Enhancement effect in 1957 (Emerson et
al., 1957). The dip in the blue (minima at 490 nm) was due to
only 40-50% efficiency of excitation energy transfer from from
the carotenoids to Chl a. The experiments were done at the
Carnegie Institution of Washington at 290 Panama Street,
Stanford, California, using a grating monochromator assembled by
Emerson and Lewis themselves. The minimum quanta of light needed
to evolve one molecule of oxygen approached 10 in these
experiments. Also shown as inserts are the photographs of late
Robert Emerson (in 1958) and the late Charleton M. Lewis (in
1996) taken by the author.
Figure 4. The quantum yield action
spectra of oxygen evolution in the diatom Navicula minima
obtained by Tanada (1951), then a doctoral student of Robert
Emerson at the University of Illinois, Urbana, Illinois, using
the Emerson-Lewis monochromator and Emerson’s perfected
Warburg manometry. Here, the existence of the "red
drop" (beyond 680 nm) was confirmed, but more importantly,
the almost 80-90% efficiency of fucoxanthin absorbing in the blue
to green region was established. Further, the minimum quantum
requirement of oxygen evolution (inverse of the maximum quantum
yield of oxygen evolution) was found to be 8. It was the same
instrument, located in the 155 Natural History Building at
Urbana, that was used later for the finding that fucoxanthin was
a major sensitizer in the then-called "short wave"
system (now PS II), whereas the Chl a absorbing beyond 680
nm was in the then-called "long wave" system (now PS I)
( see Emerson and Rabinowitch, 1960; Govindjee and Rabinowitch,
1960).
Figure 5. Absorption spectra (top
panel) and action (or excitation) spectra of Chl a
fluorescence measured at 725 nm (bottom panel ) at cryogenic
temperatures (open circles, 4K; closed circles, 77K) in Chlorella
pyrenoidosa. In addition to Chl a bands (440 nm,
670-672 nm, and 678-679 nm) and Chl b bands (477 nm and 649 nm),
a carotenoid band at 491 nm in both absorption and spectra and
action spectra of fluorescence was observed. This experiment was
done at the University of Illinois at Urbana, Illinois, by my PhD
student Frederick Yi-Tung Cho (Cho and Govindjee, 1970a).
Figure 6. Absorption spectra (top
panel) and action (or excitation) spectra of Chl a
fluorescence measured at 715 nm (bottom panel ) at cryogenic
temperatures (open circles, 4K; closed circles, 77K) in the blue
oxygenic bacterium Anacystis nidulans . In addition to Chl
a bands (440 nm, 670nm, and 679 nm) and phycobilin bands
(580 nm, 622-625 nm, 634-637 nm, and 650 nm), carotenoid bands at
around 470 nm and 502-505 nm were observed in both absorption and
action spectra of fluorescence. However, the calculated
efficiency of excitation energy transfer from the carotenoids to
Chl a was much lower than in the green algae (Cho and
Govindjee, 1970b). Unknown to the author at that time, Goedheer
(1969a,b), in Utrecht, the Netherlands, had observed similar
bands at 471 nm and 504 nm (at 77 K) in the action spectra of
several algae that were specially treated; Goedheer concluded
that these bands were from b carotene,
and that they transferred energy with 100% efficiency to Chl a,
whereas there was no energy transfer from the xanthophylls to Chl
a.
Figure 7. Action spectra of
(relative) quantum yield of Chl a fluorescence measured at 740
nm, at 77K, in grana and stroma lamellae (top panel) showing the
relative inefficiency of excitation energy transfer from the
carotenoids (the blue region) to Chl a in both the
systems. Using the methods and assumptions of Emerson and Lewis
(1943) and of Duysens (1952), the efficiency of energy transfer
to Chl a was calculated (bottom panel). The data were
suggestive of two pools of carotenoids, one absorbing in the
short wave region and transferring energy with 40-45 %
efficiency, and the other in the longwave region and transferring
energy with only 20% efficiency (after Gasanov et al., 1979). No
firm conclusions are available. Further research is necessary to
investigate the light harvesting and energy transfer capabilities
of not only carotenoids as a group, but of individual carotenoids
in PS I and II.
Figure 8. A cartoon for the
arrangement of four protein complexes in the thylakoid membrane.
Also shown, in cartoon form, is the creation of membrane
potential as the electrons are transferred from the inner side of
the membrane to the outerside of the membrane during primary
charge separation in both photosystems I and II. It has been
suggested that carotenoids, along with other pigments, sense this
membrane potential through electrochromism leading to a 515 nm
absorbance change (D A 515) (see
inset). The four major protein complexes, embedded in the
thylakoid membrane, are used for the production of the reducing
power NADPH and ATP, both needed for the fixation of CO2 and the
production of glucose. These complexes are: Photosystem II
(PSII, that oxidizes water to oxygen, reduces a plastoquinone
molecule, and releases protons in the interior of the thylakoid
membrane, and is also called water-plastoquinone
oxido-reductase); Cytochrome b/f (Cytbf) complex (that
oxidizes reduced plastoquinone, reduces a copper protein
plastocyanin (PC), and releases protons in the interior of the
thylakoid membrane, and is called plastoquinol-plastocyanin
oxido-reductase); Photosystem I (PSI, that oxidizes
reduced plastocyanin and reduces NADP+, to NADPH, and is called
plastocyanin-ferredoxin oxido-reductase); and ATP Synthase
(that uses the membrane potential and the proton gradient to
produce ATP from ADP and inorganic phosphate). The membrane
potential is formed and electron transport takes place when
photosynthesis is simultaneously powered by light absorbed in
both PS I and II leading to electron transfer from the inner side
of the thylakoid membrane to the outer side of the membrane; this
makes one side of the membrane more negative than the other. The
reaction center Chls P680 (in PS II) and P700 (in PS I) are
located on the inner side of the membrane. Photosynthesis starts
by simultaneous excitation of P680 and P700. Excited P680 (P680*)
and P700 (P700*) have energy resulting from light absorption. An
electron is transferred from P700* to Ao (another special Chl a
molecule) producing oxidized P700 (P700+) and reduced Ao (Ao-).
At about the same time, an electron is transferred from P680* to
a pheophytin (Pheo) molecule producing oxidized P680 (P680+) and
reduced Pheo (Pheo-). These are the only steps where light energy
is used to produce oxidation-reduction energy. The rest of the
reactions are energetically downhill. P700+ reduced to P700 by
receiving an electron that originates in Pheo-, and is passed on
to P700+ via the following intermediates: from Pheo- to QA (a
bound plastoquinone), to QB (another bound plastoquinone) to PQ
(freely mobile plastoquinone) to an iron sulfur protein
(FeS), to a cytochrome (Cyt f), to a freely mobile plastocyanin
(PC), and finally to P700+. On the other hand, an electron on Ao-
is passed on, ultimately, to the NADP+ via several other
intermediates (A1, a phylloquinone, Fx, FA and FB, three
iron-sulfur proteins, and Fd, ferredoxin). The missing electron
on P680+ is recovered, ultimately, from water molecules via an
amino acid tyrosine (D1-Y-161, Yz) and a manganese complex (Mn).
Four such reactions (utilizing a total of 8 photons, 4 in PSII
and 4 in PSI) are required to oxidize H2O to 102 and reduce 2
NADP+ to 2NADPH. Carotenoids are. not known to play any role in
the electron transport pathway of photosynthesis.
Figure 9. Change in
absorption spectrum of a Chlorella suspension upon
irradiation with red light. This experiment, published by Duysens
(1954), was done at the Carnegie Institute of Washington at
Stanford, California. The 515 nm change was later associated with
a shift of absorbance of some carotenoids, among other pigments,
to longer wavelengths. Further, this change, on a nanosecond time
scale, was shown to monitor the membrane potential component of
the proton motive force created in the thylakoid membrane (also
see Figure 8). Inset shows a 1963 photograph of L.N.M. Duysens.
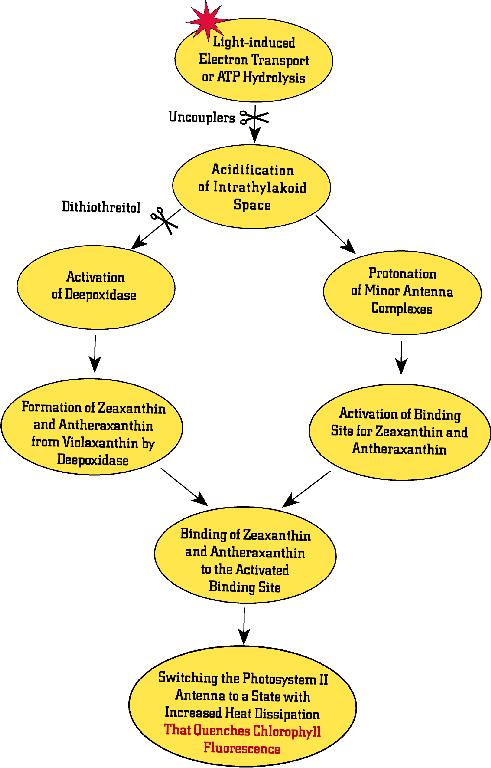
Figure 10. One of the several
schemes used to explain the mechanism of photoprotection by
xanthophyll cycle pigments. It is suggested that, at high light
intensities, when photosynthesis is saturated, there are excess
protons in the lumen of the thylakoid (see Figure 8) and these
have a double function: (1) they protonate minor antenna
complexes of PS II leading to the activation of a binding site
for xanthophylls (see right side of the diagram); and (2) they
activate de-epoxidase allowing the conversion of violaxanthin
into antheraxanthin and then into zeaxanthin (see left side of
the diagram). The two events together then lead to the quenching
of Chl a fluorescence through dissipation of energy as heat. The
scheme shown here was modified from Gilmore and Govindjee (1998).
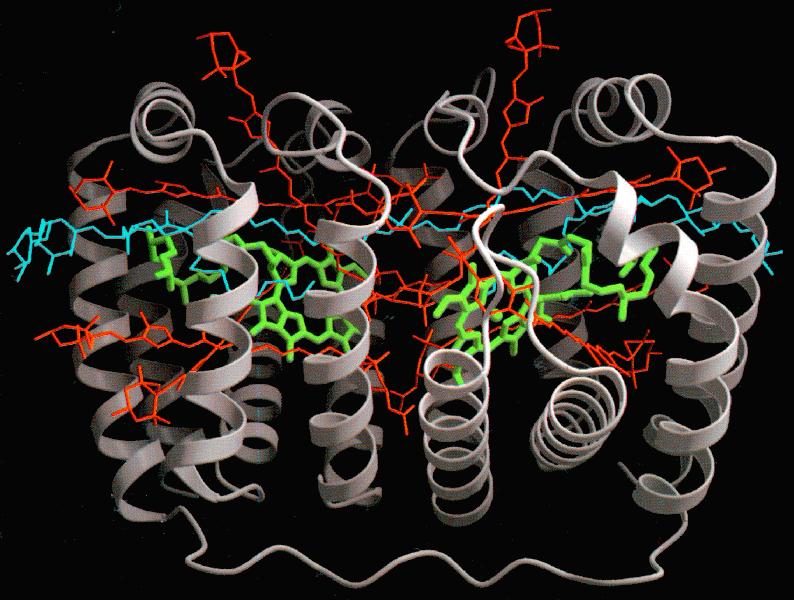
Figure 11. A 2 Å resolution
structure of a peridinin Chl a complex of a dinoflagellate. The
eight peridinin molecules are shown in red, whereas the two Chls
are in green. A lipid molecule is shown in blue, and two proteins
are shown in grey The proximity of peridinin molecules to each
other and to Chl a molecules explains the efficient
excitation energy transfer from peridinin to Chl a, observed
by Haxo et al. (1976). The diagram is reproduced from Hoffman
et al. (1996). See Hoffman et al. (1996) for Fig. 11
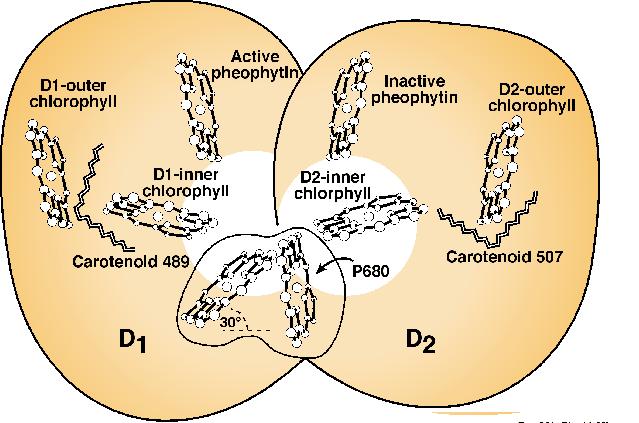
Figure 12. A diagrammatic model of the arrangement of two carotenes (labeled as carotenoid 489 and carotenoid 507) in the reaction center molecule of PS II. P680 is the reaction center Chl a dimer, whereas D1 and D2 are the two proteins where the chromophores are housed. The scheme shown here was modified and adapted from Mimuro et al. (1995). For a more complete model of Photosystem II reaction center and a different view of the arrangement of carotenoids, see Xiong et al. (1998).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}