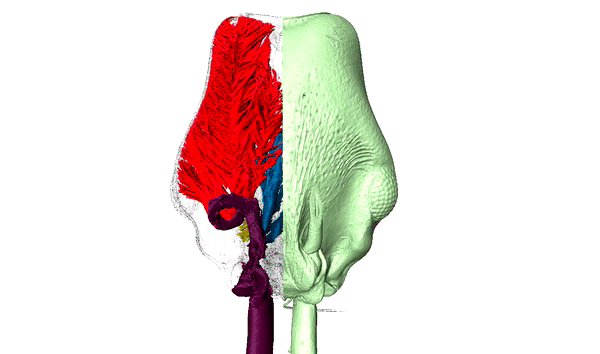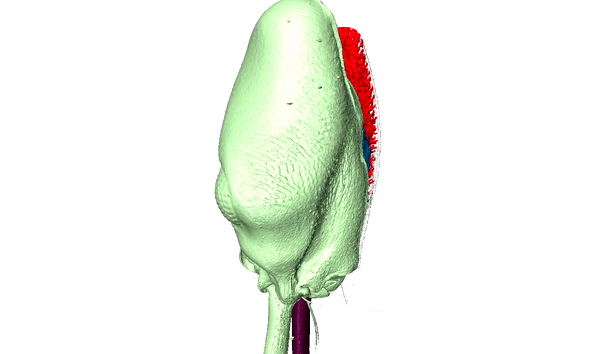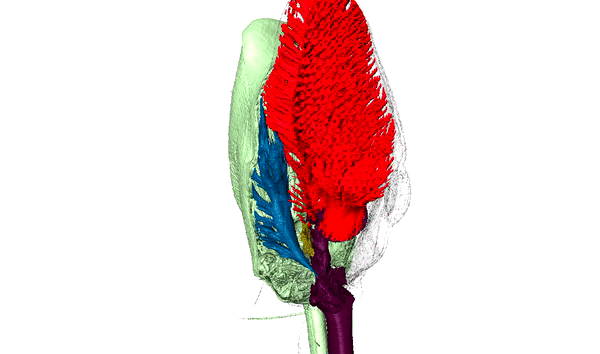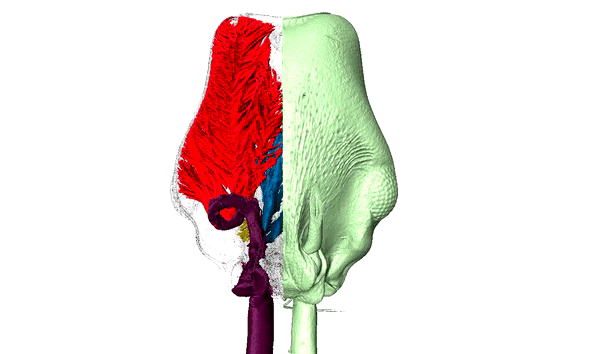
Ants are among the most diverse and successful organisms in terrestrial ecosystems. In addition to having over 15,000 described species, this single family of insects can make up over 1/4 of the animal biomass in the tropics where they act as scavengers, top-predators, herbivores, and engage in a variety of mutualisms. Our research capitalizes on the developmental and ecological flexibility of ants to investigate how polymorphism and specialization within complex societies contribute to their ecological success.
Suarez Lab
Ant ecology and behavior at the University of Illinois