Excitation Energy Transfer and Energy Migration : Some Basics and Background
(by Govindjee; a portion of the text has been modified from Rabinowitch and Govindjee, 1969, Photosynthesis, John Wiley & Sons)
If there are two molecules in close proximity, one with an absorption band at a wavelength shifted to the longer wavelengths than another, light energy absorbed by the one absorbing at the shorter wavelength is usually transferred to the one that absorbs at the longer wavelengths. That is, one molecule acts as a donor of excitation energy, and the other as an acceptor of this energy. In photosynthetic systems, the acceptor is a chlorophyll a molecule, and the donor could be chlorophyll b (in green plants), phycocyanin (in cyanobacteria), or fucoxanthin (in diatoms and brown algae), among other pigments (chromophores). This transfer probably takes place by a resonance mechanism, similar to the one familiar from acoustic experience, but properly describable only in terms of quantum mechanics. There are reasons to believe that most chlorophyll a molecules also do not participate directly in the primary photochemical process in photosynthesis, but transfer their excitation energy to the few chlorophyll a molecules directly associated with the reaction centers that lead to the primary charge separation. Excitation energy is in the form of a couple, an electron that is in the excited state of the molecule with the "hole" it has left behind in the ground state. What is transferred is the "excited electron and the hole". We shall call this entity an "exciton". Thus, we can talk about exciton transfer.
We shall deal here with energy transfer between different pigments ("heterogeneous" transfer), as well as with transfer between identical molecules ("homogeneous" transfer). The latter can be repeated many times, giving rise to energy migration. Direct evidence of energy transfer between different pigments is provided by sensitized fluorescence. Light quanta absorbed by molecules of one pigment (for example, chlorophyll b) are transferred to molecules of another pigment (for example, chlorophyll a). When the first pigment is excited, only fluorescence of the second is observed. This phenomenon, of sensitized fluorescence, is well known from studies on gases and solutions. The occurrence of heterogeneous energy transfer from various pigments to chlorophyll a is inferred from the action spectra (or "excitation spectra") of chlorophyll a fluorescence in vivo.
Action (or Excitation) Spectra of Fluorescence
If we plot, as a function of wavelength of the exciting light, the yield (per incident number of photons) of chlorophyll a fluorescence (measured in the long-wave region, where fluorescence is not reabsorbed) in higher plants or green algae, we obtain curves that approximately parallel the action spectra of photosynthesis, i.e., the rate of photosynthesis as a function of wavelength of light. Since absorption by chlorophyll b is maximal at 480 and 650 nm, presence of peaks at these wavelengths shows that chlorophyll b, an accessory pigment, sensitizes photosynthesis by transferring its excitation energy to chlorophyll a.
In the case of "ideal" resonance, such as that exists between atoms of sodium in sodium vapor, or even molecules of chlorophyll a within a pigment-protein complex, the size of quanta absorbed by one molecule equals precisely that absorbed by the others. When such identical atoms or molecules are extremely close together, as in a dense vapor, concentrated solution, or a crystal, the interaction forces between adjoining molecules may cause the excitation quantum to essentially become a communal property of all of them.
The mechanism of resonance transfer of energy between unlike molecules, without (or, with only partially) overlapping absorption bands, was analyzed in 1948 by the German physicist Theodor Förster.
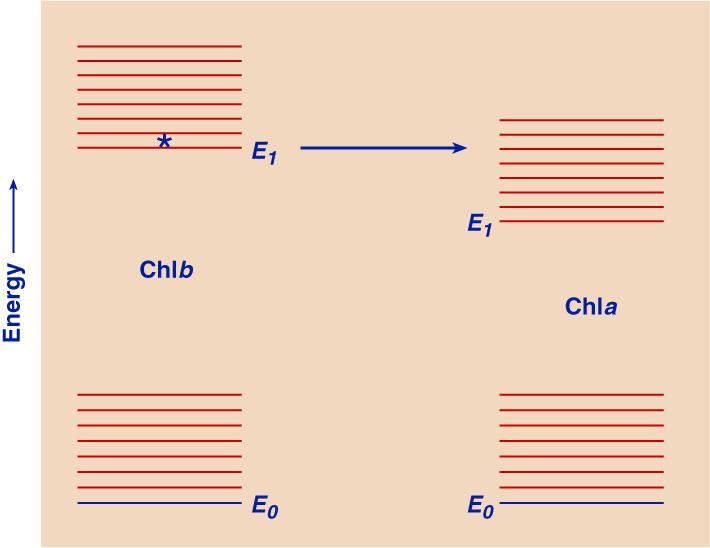
FIG. 1.
Energy level diagrams of Chl b and Chl a, indicating energy transfer from the lowest singlet excited state of Chl b to that of Chl a. This transfer has a high probability because the process is "down-hill".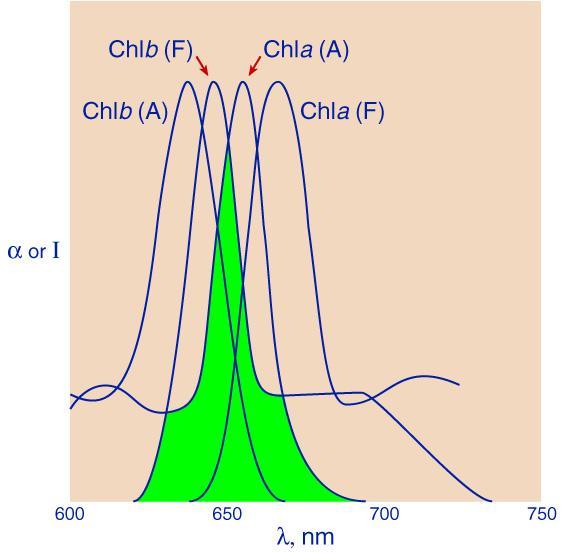
FIG. 2.
Overlapping of absorption band (A) of Chl a and fluorescence band (F) of Chl b (shaded area), and of the absorption bands of the two pigments.The absorbed quantum belongs, at first, to one pigment molecule only. However, in the process of vibrational energy dissipation (internal conversion) in the excited electronic state of the "donor" pigment (say Chl b), states are reached that are in resonance with certain (strongly vibrating) states of the "acceptor" chlorophyll a (FIG. 1). This resonance is what makes energy transfer possible. According to Förster, there are three parameters that control this excitation energy (or exciton) transfer. (1) One measure of probability of such a transfer is this overlapping of the fluorescence band of the donor and the absorption band of the acceptor, indicated by shading in FIG. 2, the so-called Förster's overlap integral. (2) A second measure is related to the distance between the two molecules. The interaction between molecules with overlapping absorption bands is a dipole-dipole interaction. The energy of such an interaction is proportional to r-3, where r is the distance between the centers of the two dipoles (presumed to be large, compared to the distance between the two poles in each dipole). The interaction** between molecules in which the fluorescence band of one overlaps the absorption band of the other, caused (as previously mentioned) by resonance is, on the other hand, a "second order" effect; as such, it is proportional to r-6. (3) The third measure is the so-called "orientation factor" related to the orientation of the dipoles of the donor and the acceptor molecules.
-------------------------------------------------------------------------------------------------
**
The hierarchy of potential energies, E (and of interaction forces, F, which are the derivatives of potential energies with respect to distance, F = dE/dr), begins with the Coulomb interaction between two free charges ("monopoles"). In this case, E is proportional to r-1, and F to r-2. The interaction energy between a monopole and a permanent dipole is proportional to r-2, and the force between them, to r-3. The energy between two permanent dipoles is proportional to r-2 and the force between them to r-4. The interactions involving an induced (rather than a permanent) dipole (that is, a dipole formed in a nonpolar molecule under the influence of a monopole, or of another dipole) are "second order" effects (because the moment of the induced dipole is itself proportional to the interaction force). The corresponding energy is proportional to (r-2)2, or r-4 in the case of monopole-induced dipole interaction, while the interaction energy of two mutually induced dipoles is proportional to (r-3) 2 or r-6. This is the type of interaction that quantum mechanics suggests exists between unlike molecules resonating in the above described "delayed" way. Current research in the 1990s in several laboratories indicate that excitation energy transfer in many photosynthetic systems may not follow Förster theory although they involve coulombic interactions. Also, they may involve multipoles instead of dipoles. This will not be discussed here. The reader may look up the work of Graham Flemming and of Klaus Schulten, among others..------------------------------------------------------------------------------------------
Förster's calculations suggested that, if the overlapping between the fluorescence band of an excited "donor" molecule and the absorption band of a nonexcited "acceptor" molecule is substantial (as in FIG. 2) the time needed for the excitation energy transfer may become equal to the natural lifetime of the excited state when the distance between two molecules still is considerably larger than the sum of their "kinetic radii (that is, radii determined by gas-kinetic methods). In other words the probability of energy transfer can reach 50% long before the two molecules actually touch each other. The calculated (and actually observed) "critical distances" over which the transfer probability equals 50%, are of the order of 5 nm. In a chloroplast, the distance between different pigment molecules much less than 5 nm, so that the probability of energy transfer must be quite high.
Because of the Stokes shift (see the earlier discussion on the Frank-Condon principle), the fluorescence band of a pigment absorbing at the shorter wavelengths often overlaps the absorption band of a pigment absorbing at somewhat longer waves, but not vice versa (FIG. 2). Consequently, energy transfer usually can go in one direction only. For example, energy transfer goes from pigments absorbing in the blue region of the spectrum (carotenoids,e.g., fucoxanthin) to Chlorophyll a in diatoms and brown algae.. In red algae, this transfer is from pigments absorbing in the green (phycoerythrin) to those absorbing in the orange (phycocyanin); and ultimately to those absorbing in the red ( first, allophycocyanin, and then chlorophyll a). In each transfer, discussed above, some electronic energy is converted into vibrational energy (and then dissipated into thermal energy).
We shall now consider experimental evidence of this kind of excitation transfer in vivo. The first relevant observation was made in 1943 by H. J. Dutton, W. M. Manning, and B. M. Duggar at the University of Wisconsin. They measured the yield of fluorescence of chlorophyll a in a diatom, using monochromatic excitation, and found that this yield was almost the same whether the incident light was absorbed by chlorophyll a, or by fucoxanthin. This suggested that most light quanta taken up by fucoxanthin were transferred, by resonance, to chlorophyll a. Subsequently, C. S. French and co-workers in California, and L. N. M.Duysens in Utrecht in the Netherlands, found that a similar transfer goes on, with varying effectiveness, between all accessory pigments in photosynthesizing cells and chlorophyll a as the final energy acceptor. Least efficient is the transfer from the some of the yellow carotenoids; only 20-50 percent of the quanta absorbed by carotenoids find their way to chlorophyll a. Much more effective--of the order of 80–90%--is the transfer from phycoerythrin to chlorophyll a in red algae. At the University of illinois, first S. Brody and then G. Tomita, working with Eugene Rabinowitch, determined directly, for the first time, the time lag of emission of chlorophyll a fluorescence after absorption in phycoerythrin (or phycocyanin) as compared to its emission after absorption in chlorophyll a itself. The transfer times were found to lie between 0.3 and 0.5 nanoseconds (ns). In other words, they were considerably shorter than the average emission times of phycoerythrin fluorescence (7.1 ns) or phycocyanin fluorescence (1.8 ns). Consequently, only a small amount of phycoerythrin or phycocyanin fluorescence is emitted in competition with energy transfer from these pigments to chlorophyll a. SEE a figure from Yamazaki et al. (1984) showing the cascade of energy transfer in red algae (handout on March 25, 1999).
Still more efficient is the transfer from chlorophyll b to chlorophyll a- close to 100 percent; the same must be true for transfers from one Chl a molecule to another in the same pigment-protein complex. Chlorophyll a molecules are differently bound to the proteins, and, thus, there are several different spectral forms of chlorophyll a in vivo. They are Chl a 670, Chl a 680, Chl a 690, etc (the numbers indicating the wavelenghths, in nanometers, of the peaks of their first singlet excited states). Excitation energy transfer among them must occur by a mechanism similar to that from Chl b to Chl a, downhill energywise. In the case of bacteriochlorophyll in photosynthetic bacteria, the light quanta absorbed by two forms of this pigment (those with absorption bands at 800 nm and 850 nm) are transferred to the third one, with the absorption band at about 890 nm, and practically all fluorescence is emitted from the latter. A unique case of "uphill" energy transfer exists in the photosynthetic bacterium Rhodopseudomonas viridis where the reaction center bacteriochlophyll b has its first singlet excited state at much shorter wavelengths (980 nm) than the large number of bacteriochlophyll b molecules that do most of the absorbing (1,050 nm).
The study of sensitized fluorescence is an elegant method to follow the fate of excitation energy in multipigment systems in plant and bacterial cell as well as in pigment-protein complexes isolated from them. The main conclusion from this study is that transfer of excitation energy (or excitons) occurs from the accessory pigments to the reaction center chlorophyll a or bacteriochlorophyll depending upon the system we are investigating. . Although these transfers are rather fast (femtoseconds to picoseconds), some energy is also lost as fluorescence, or as heat, depending upon the value of the various rate constants of the various de-excitation pathways.
Excitation energy transfer between identical pigment molecules, particularly between chlorophyll a molecules in "photosynthetic units" cannot be determined by the "sensitized fluorescence" technique, i.e., observation of acceptor fluorescence when you excite the donor molecule. However, one observes quenching of fluorescence, i.e., reduced fluorescence when excitation energy transfer is a major event leading to photochemistry. Also, if the molecules are arranged randomly, excitation with polarized light leads to depolarization of fluorescence if there is excitation energy migration.
The consequences of resonance interaction depend on its strength. In one extreme case-that of "weak coupling"-a proper approximation may be the picture of excitation energy migrating by a sequence of jumps" between adjoining molecules-a kind of "random walk" of the excitation quantum. On the opposite end, that of "strong coupling," one must instead imagine a simultaneous, "communal" excitation of all pigment molecules involved in the exchange (forming, as it were, a single supermolecule"). The latter case is well known from the study of certain dye polymers, in which strands of several hundred molecules behave like a single giant molecule. "Weak" and "strong" interaction are defined by comparison of the interaction energy with the vibrational energy in the individual molecules. If the interaction energy is smaller than the vibrational energy in the individual molecules, the absorption process occurs, in the first approximation, within an individual molecule, and the intramolecular vibrations (whose excitation converts an absorption line into an absorption band. However, when the interaction energy is much larger than the vibrational energy, the excitation exchange frequency, n e, is (according to the basic law of quantum mechanisms, E = hn ) higher than the vibration frequency, so that numerous energy transfers can take place during a single vibration. Under these conditions, electronic excitation becomes a "communal" phenomenon, and intramolecular vibrations are uncoupled from electronic excitation, with consequent far-reaching change in the shape of the absorption hand.
In the case of chlorophyll a in vivo, the apparent similarity of the absorption spectrum of the pigment in vivo with its spectrum in solution (in which absorption occurs in isolated molecules), suggests that conditions in photosynthetic units may not be those of "strong interaction" in which individual molecules are combined into a supermolecule. Rather, in most photosynthetic systems, the situation may approach the case of "weak interaction," in which excitation moves around from molecule to molecule in a random walk.
The random walk of excitation in the photosynthetic unit energy ends when the quantum is either reemitted as fluorescence by one of the molecules it visits on the way, or is dissipated by internal conversion in one of them, or reaches a "trap" (a spot where its energy is used to bring about a chemical reaction, also called the reaction center.) We believe this is what often happens in the photosynthetic units in chloroplasts; but nobody has been able to devise an experiment directly demonstrating the random walk of the quantum in them. An indirect evidence of excitation migration is almost complete depolarization of chlorophyll a fluorescence in vivo when excitation is by polarized light.
Simple arguments can be adduced in favor of migration of the excitation quantum as an indispensable step in photosynthesis. In order to utilize efficiently solar radiation, plant cells must strongly absorb it. A leaf, containing a few layers of green cells, in fact absorbs red and blue-violet light almost completely. Even a single cell of a green alga Chlorella, about 5 X 10-4 cm thick, absorbs up to 60% of incident light in the maximum of the red absorption band--this is why it appears distinctly green under the microscope. To achieve such absorption, it is not enough for the cells to contain a strongly absorbing organic pigments, such as chlorophyll (or a phycobilin), they must be present in large amounts. The most intensely colored pigment cannot absorb more quanta than its absorption coefficient permits***
---------------------------------------------------------------------------------------------------------
***
A molar absorption coefficient of 105 means that 6 x 1023 molecules have a total "opaque" cross section of l05 X 102 cm. (The factor 102 appears because concentration is measured in moles/liter.) This means 107/6 X 1023 = 1.6 x 10-17 cm2, or 15 A2, per molecule-which is close to the "kinetic" cross section of a medium-sized organic molecule, such as chlorophyll, as determined, for example, from its diffusion coefficient. In other words, pigment molecules are practically "black" to light in the peak of their absorption band.------------------------------------------------------------------------------------------------------------
The highest absorption coefficients of organic pigments are of the order of 105 Typical green plant cells have linear dimensions of the order of 10-3 cm. To produce a 50% absorption in the absorption band maximum (I = 0.5I0), the pigment concentration in such a cell must be of the order of 3 X 10-3 mole/liter, as the following simple application of Beer's law proves. If I is equal to 0.5I0, we can write (a stands for extinction coefficient of the pigment; c stands for concentration of the pigment; and d stands for the pathlength):
log (I0/I) = log 2 = 0.3 = a cd = 105c10-3; or,
c = 3 X 10-3 mole/l (Eqn. 1)
Actually, fully green leaves do contain up to five percent of chlorophyll a in relation to dry weight, corresponding to a cellular concentration of the order of 10-2 mole per liter.
The purpose of this estimate was to make clear that by the very nature of the task placed before photosynthesizing cells, they must be densely packed with pigment molecules. Now, having absorbed the light quanta, the cell must use the absorbed energy to set in motion chains of enzymatic reactions, by which the unstable primary photochemical products arc converted into the final products. For this, at least a dozen, if not more, specific enzymes arc needed. An enzyme molecule is a protein with a molecular weight (and thus also space requirement) a hundred or thousand times larger than that of a chlorophyll molecule. There is not enough space in the cell for a separate "enzymatic assortment" to be assigned to each pigment molecule! A large number of pigment molecules simply must share a common enzymatic "conveyor belt."
Fortunately, the catalytic capacity of enzymes is quite sufficient for this purpose. It is easy to calculate, from Beer's law, that a single pigment molecule, with an absorption coefficient a will absorb, in a second, out of a light flux Ihn , (measured in number of quanta falling each second upon one cm2), a number, n, of quanta determined by the following equation:
n = 4 X 10-21 a Ihv (Equation 2)
The strongest flux to which plants are exposed in nature is that of full sunlight at noon, which corresponds to 1016 - 1017 (visible) quanta per sec per cm2. This means according to Equation 2 that each pigment molecule will absorb, under natural conditions, only a few quanta (n = 4 - 40) each second. This is very little by standards of enzymatic efficiency; many enzyme molecules can handle tens of thousands of substrate molecules per second. One "enzymatic conveyor belt" is, therefore, quite sufficient to handle the photochemical production of hundreds, if not thousands, of pigment molecules.
It has been known for about 37 years that chloroplasts contain hundreds of chlorophyll molecules per "reaction center chlorophyll" molecules. A mechanism is required by which light quanta absorbed by several hundred chlorophyll molecules can be put to work on a single enzymatic conveyor belt–a system of "messengers" connecting the many pigment molecules in a unit with a single reaction center. These messengers could conceivably have been material particles, for example, the primary oxidation or reduction products, diffusing from the many pigment molecule where the quantum had generated them, to a single "reception point" of the enzyme system. A more compact, safer, and faster mechanism is, however, physically possible: instead of a messenger with a post office letter, the communication can be by Electronic mail (e-mail)! This is what resonance energy migration amounts to. In it, only excitation energy, in the form of "excitons"-in other words, only the light electrons and not the heavy atoms or molecules, needs to move.
It is the nature of a quantum phenomenon that the energy of a migrating "exciton" is not dissipated during migration, but kept in one piece. The course of migration can be represented by a spreading wave, but the whole energy of the "exciton" can be found, at a given moment, in one or the other of the many resonating molecules. The wave describes merely the spread of the probability of finding the quantum in different locations. One can visualize the exciton as consisting of an excited electron an "electron hole" left in the ground state of an atom or molecule. In "random walk" energy-the case we consider here, the electron and the hole move together, always remaining in the same atom or molecule (FIGURE 3). In other words, this type of migration involves no separation of positive and negative charges.
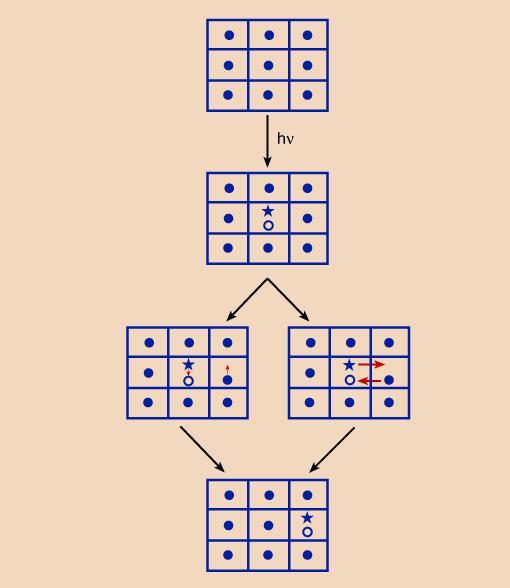
FIG. 3
Two mechanisms of exciton migration in a crystal lattice. Left: Heller-Marcus mechanism; right: Wannier mechanism. Dots represent unexcited molecules, asterisks, excited electrons, and circles, positive cores left after excitation.The random walk picture of energy migration reminded Eugene Rabinowitch of a pinball machine, into which a steel ball had been shot; and the different bulbs lighted up one after another, as the ball ran around on the board, until it either fell to the bottom (which corresponds to reemission of the quantum by fluorescence), or fell into a "pay hole," that is,. its energy was utilized for a photochemical process. It is worth mentioning here that resonance energy migration does not in itself affect the natural lifetime of excitation. However, in practice, the random walk of the "exciton" involves risks. A migrating quantum has a greater chance of getting into a loss situation than a stationary one, and this may lead to its premature "death". More specifically, some molecules in the array may be engaged in chemical interactions. This can create a "trap" (or "sink") into which the migrating quantum stumbles. In other words, resonance energy migration can lead to the abbreviation of the lifetime of the exciton, and thus to the quenching of fluorescence. In photosynthesis, the trap may be identical with the "reaction center," where the migrating quantum is caught and put to useful work. However, other traps may be formed that may lead to heat loss as has been recently suggested when a carotenoid zeaxanthin binds to specialized minor chlorophyll-protein complexes. In fact, it has been suggested that this is the mechanism by which plants protect themselves against damage by excess light.
Resonance migration accounts, at least in part, for the well-known phenomenon of concentration quenching of fluorescence. It is known that strong solutions of fluorescent pigments fluoresce weaker than the more dilute ones. An even more sensitive index of resonance energy migration is depolarization of fluorescence--weakening or disappearance of polarization normally present in fluorescence excited by polarized light, as mentioned earlier. If fluorescence is excited by plane-polarized light, that is, light that vibrates (and causes, upon absorption, the electrons in the molecule to vibrate) preferentially in a certain plane, the emitted fluorescence also is polarized. This is so because in the interval between absorbing a quantum and reemitting it, the molecule does not have enough time to forget the orientation it has had at the time of absorption. But if the energy quantum undergoes, between absorption and emission, a series of resonance transfers, each molecule in the resonance chain will be oriented somewhat differently, and after a few transfers, the original preference for a certain direction will be lost. It has been observed, for example, that the fluorescence of phycobilins (phycoerythrin and phycocyanin included), excited by polarized light, is completely depolarized. This can be considered as evidence that a quantum absorbed by one of many phycohilin molecules contained in the pigment-protein complex, moves, by resonance, from one of them to another and becomes completely "disoriented" by the time of reemission.
SIMULTANEOUS MIGRATION OF ELECTRONS AND HOLES BETWEEN TWO MOLECULES
There is another mechanism of energy migration. It is called the Dexter mechanism.. When the molecules are almost touching each other and the interaction energy between them is high, there is the possibility that (see the right arm of figure 3, called the Wannier mechanism there) as the excited electron is transferred to (say from molecule A) the empty higher energy level of the next molecule (say, molecule B), the electron in the ground state of the molecule is simultaneously moved to fill the "electron hole" of molecule A. The net result is the same as in the Foerster transfer, but here the electrons physically move.
FURTHER READING:
[1] Handout of March 25, 1999 labeled "Light Absorption and Energy Transfer"
[2] R. van Grondelle and J. Amesz (1986) Excitation Energy Transfer in Photosynthetic Systems. In:Light Emission by Plants and Bacteria (Edited by Govindjee, J.Amesz and D.C. Fork), pp. 191-223, Academic Press, Orlando.
[3] J. Lakowicz (1986) Principles of Fluorescence Spectroscopy. Plenum Press, New York (see chapters 1, and 10).
(4) Govindjee (1999) in Frank, H et al. (ed.) Photochemistry of Carotenoids, Kluwer Publishers, Dordrecht, in press; see the Link elsewhere.